Lake Ecosystem Research New Zealand (LERNZ) is a collaborative effort between scientists from the University of Waikato and partner organisations, committed to preserving and restoring aquatic ecosystems. Rooted in limnological principles, our research addresses ecosystem structure and function, scale, connectivity, and processes such as disturbance, flows, and resilience. By employing methodologies like remote sensing, fieldwork, experiments, and modelling, we advance our understanding and management of freshwater ecosystems. Driven by the imperative to mitigate the impacts of climate change and anthropogenic pressures on aquatic ecosystems, our research investigates topics ranging from biological invasions and habitat restoration to water quality management. We collaborate with partners across iwi, regional councils, the Department of Conservation, the Ministry for the Environment, and other research institutes.
Successful Endeavour Smart Idea: An Ecosystem Modelling Platform to Assist New Zealand Lake Management
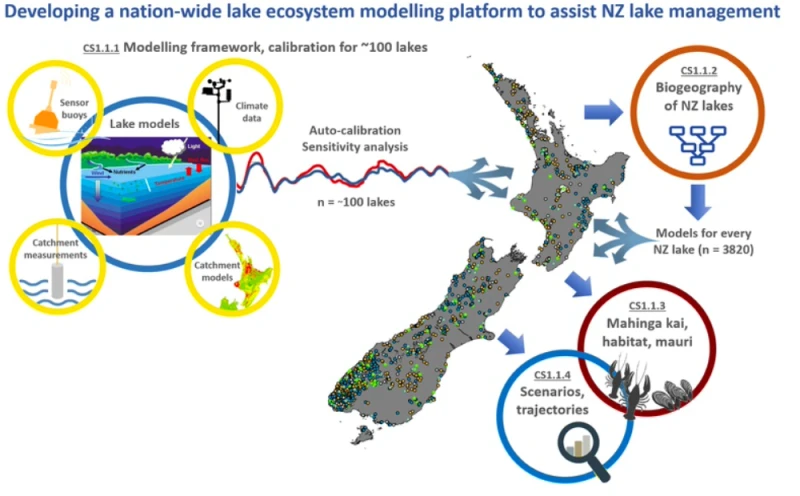
LERNZ researchers Deniz Özkundakci, Chris McBride, Moritz Lehmann, Annika Hinze, Mat Allan and Ian Kusabs received $1m over the three-year project to build a platform modelling the ecosystems of New Zealand's 3820 lakes. Using supercomputing for autocalibration and ensemble simulation, leverage already available data, and exploit satellite remote sensed water quality information. The project team is supported by top national and international scientists to ensure the science is cutting edge and applicable outside New Zealand.
Research lead: Deniz Özkundakci